11.4.1. Elméleti alapok
A különböző élőlény populációk terepi vizsgálata során mindig szembesülünk a különböző fajok eltérő szaporodási stratégiáival. A megbolygatott élőhelyeken agresszíven terjedő gyomnövények (pl. parlagfű) szinte egyik napról a másikra alkotnak jelentős állományokat és az emberi tájrombolás (pl. taposás, égetés, építkezés) vonalai mentén igyekeznek előretörni a még természetközeli társulásokba, felborítva az ezekben, több évszázad alatt kialakult kényes biológiai egyensúlyt. Hasonló módon viselkednek azok az idegen tájakról behurcolt állati kártevők is, mint egyes lepkék hernyói, amelyek akár egész erdőrészeket tarolnak le rágásukkal. Mielőtt abba a hibába esnénk, hogy a külföldi országokat hibáztatnánk e jelenség okán, meg kell jegyeznem, hogy a hazánkban természetes fajoknak számító növények és állatok Amerikába behurcolva hasonló károkat okoznak. Ugyanakkor a stabil természetes életközösségekben élő fajok, a környezeti tényezők viszonylagos állandósága mellett nem szaporodnak túl, egyedszámuk nem változik jelentősen az idő előrehaladtával. Ahhoz, hogy megértsük a két eltérő szaporodási stratégia alapját, meg kell vizsgálnunk a populációnövekedés mozgatórugóit. Ebben és a különböző stratégiák modellezésében a számítógép lesz segítségünkre.
A populációnövekedés két legegyszerűbb formája az exponenciális (korlátok nélküli) és a logisztikus (korlátos) növekedés. Exponenciális növekedés esetén a populáció növekedési üteme (ha az egyedek átlagosan ugyanannyi utódot hoznak létre egy időintervallumban) a populációban már jelenlevő szervezetek számának egyszerű többszöröse. Tehát, ha egy élőlény átlagosan két új utódot ad át a következő nemzedéknek, akkor két élőlény négyet, tíz élőlény huszat és így tovább. Ha az egyedek szaporodási üteme állandó, akkor a tíz egyedből álló populáció tízszer gyorsabban növekszik, mint az egyetlen élőlényből álló.
Abban az esetben, ha az egyedek csak bizonyos évszakokban szaporodnak és az egyes nemzedékek nem fednek át (minden egyes nemzedék kicserélődik a következőre, azaz a szülő egyedek elpusztulnak, csak az utódok élnek tovább), akkor átlagosan két utód esetén a populáció minden nemzedékben megkétszereződik. Általánosan megfogalmazva, ha R a nemzedék újratermelődési rátája (példánkban R=2), N a populáció mérete (az egyedek száma a populációban), N0 a kiindulási egyedszám, és t az elmúlt generációk száma, akkor az aktuális egyedszám a következő képlettel határozható meg:
N = Rt N0
Az exponenciális növekedés másik szélsőséges esete az, amikor a populációban folyamatos a szaporodás. Ezt a folyamatot a következő egyenlet írja le.
dN/dt = rN = (b0-d0)N
N a populáció egyedszáma adott pillanatban
t az idő tetszőleges egységben mérve
r állandó a belső szaporodási ráta
b0 az egy egyedre eső átlagos utódszám egy időegység alatt
d0 az egy egyedre eső átlagos halálozási szám időegység alatt
A modell szerint a populáció növekedési ütemét egy állandónak és a már jelenlévő egyedek számának szorzata adja meg. Az állandó a populáció belső szaporodási rátája (r), ami egyenlő a b0-d0 értékével. Az r értéke a valóságban persze lehet negatív és változhat is, hiszen a születési és halálozási arányok sem állandóak még ugyanabban a környezetben sem, N változásával ezek is változnak. Mindezek ellenére vannak olyan körülmények, amelyek között, legalábbis egy ideig a populáció úgy növekedik, mintha az r=(b0-d0) állandó volna. Ez olyankor fordulhat elő, amikor a populációméret jóval a környezet eltartó képessége alatt van. Elvben minden populáció rendelkezik egy maximális belső szaporodási rátával, amely egy optimális környezetben valósulhatna meg (bőségesen rendelkezésre álló hely és erőforrások, ragadozók és vetélytársak hiánya). Ennél jóval alacsonyabb a megvalósult belső szaporodási ráta.
A populációnövekedés csak különleges körülmények között és rövid ideig felel meg az exponenciális egyenletnek. Ha ugyanis ez nem így lenne, akkor például az emberi faj, amely egyike a leglassabban szaporodó szervezeteknek, exponenciális növekedése esetén 5000 év alatt elérné a Világegyetem tömegét és majdnem fénysebességgel tágulna kifelé.
Hosszú periódusokban a dN/dt majd minden populációban 0-hoz közel álló érték, ami azt jelenti, hogy a populáció mérete egy átlagos érték körül ingadozik. Az úgynevezett logisztikus növekedési görbe az egyik lehetséges modell, amely alkalmazásával az exponenciálisan növekedő populáció megközelíti a korlátját. Ezt a határt nevezzük a környezet eltartó képességének, és K-val jelöljük. A logisztikus egyenlet (Verhulst-Pearl féle logisztikus egyenlet) az alábbi formában írható fel:
dN/dt = rN(K-N )/K
Az egyenlet megalkotásakor feltételezzük, hogy az exponenciális egyenletben szereplő b0 és d0 értékek nem függetlenek a populáció méretétől. Lehetséges, hogy bizonyos alacsony N értékeknél b és d-re nem hat az egyedszám (N) változása, ami megváltoztatja a populáció sűrűségét (egységnyi területre jutó egyedszámot). A sűrűség függő hatások a populációsűrűség függvényében megváltoztatják a születési és a halálozási ráta értékét. Ilyen hatás például a versengés a populáció egyedei között, kémiai környezet kiválasztás vagy anyagcseretermékek miatti megváltozása, a táplálékhiány, a paraziták és a ragadozók támadásainak gyakoribbá (vagy ritkábbá) válása, az elvándorlás. A népesség növekedésével a halálozási arány valószínűleg megnő, a születések aránya viszont valószínűleg lecsökken. Ha ezeket a feltételezéseket be akarjuk építeni a modellünkbe, akkor figyelembe kell vennünk b0 és d0 egyedszámtól (N) való függését a következőképpen:
b = b0 -kbN és d = d0+kdN
ahol kb a születés aránya csökkenésének meredeksége és kd a halálozási arány növekedésének meredeksége. Ezek után az egyenletünket a következőképpen írhatjuk fel:
dN/dt = ((b0 -kbN)-(d0+kdN))N
Ha az időegység alatti egyedenkénti átlagos születési (b) és halálozási ráta (d) egyenlő (b=d), akkor az egyedszám megmaradhat egy egyensúlyi értéken és ilyenkor igaz, hogy:
b0 -kbN = d0+kdN
N = (b0 -d0)/ (kb+kd)
N-nek ez az értéke a környezet eltartóképessége s általában K-val jelölik. K-nál nagyobb N esetén a populáció mérete csökkeni fog, K-nál kisebb N esetén a népesség növekszik. A K tehát olyan egyensúlyi populációméret, amelyet időben közelít minden populáció.
Egyesítve a jelöléseket:
K = (b0 -d0)/ (kb+kd) és r = b0 -d0
Behelyettesítve ezeket a logisztikus egyenletbe a következő kifejezést kapjuk::
dN/dt = rN(K-N/K) = rN(1+N/K)
Már korábban említést tettünk a sűrűségfüggő hatásokról, amelyek általában döntő szerepet játszanak a populációméret szabályozásában. Ugyanakkor léteznek olyan környezeti hatások is, amelyek anélkül változtatják meg a születési és halálozási arányt, hogy a populációsűrűség befolyásolná hatásukat. Ilyen lehet például egy környezeti katasztrófa hatása pl. bozóttűz, áradás, vulkáni hamueső. Ezek az úgynevezett sűrűségtől független hatások. Fontos belátnunk azt, hogy azok a populációk, amelyeknek a növekedését kizárólag a sűrűségtől független hatások szabályozzák, hamar kipusztulnak, mert ha egy káros hatás után nincs sűrűségfüggő szabályozásuk, ami felgyorsíthatná a növekedésüket, akkor a populációméret könnyen elérheti a zérust. Ezért feltételezhetjük, hogy a legtöbb létező populációban működik valamiféle sűrűségfüggő szabályozás.
r és K stratégia
Egy populációban az r és a K értékét a populáció genetikai összetétele határozza meg, amely értékek az evolúció során változhatnak. A kiszámíthatatlan (állandóan változó vagy csak rövid ideig létező) környezetben (pl. folyópartok iszapos részein, erdők füves tisztásain, a tundra élőhelyein élő fajok akkor sikeresek, ha gyorsan felfedezik az élőhelyet, gyors szaporodásuk eredményeként hamar benépesítik és felélik forrásait, mielőtt más versengő fajok is megtelepednének itt, majd miután kiaknázták az élőhely lehetőségeit szétszóródva új életteret keresnek. Az ilyen fajok r értéke nagy, ezeket az élőlényeket „r-stratégistáknak” is nevezik. A magas szaporodási ráta (r) lehetővé teszi sok utód élete árán, hogy egy részük újra megtalálja azokat az üres élőhelyeket, ahol az életciklus újra kezdődhet.
A „K-stratégista” fajok állandóbb környezetben élnek. Populációik egyedszáma a környezet eltartóképességének (K) közelében van. Az itt élő fajok számára a gyors szaporodással szemben előnyösebb a környezethez való alkalmazkodás, a versengésre való képesség, a környezet újabb részeinek megszerzése, megtartása, energiáinak kiaknázása. Egy stabil környezetben élő populáció számára nem jelent előnyt a magasabb r érték, sőt a szükségesnél nagyobb energia, amit a szaporodásba fektetnek, csökkentheti az egyedek túlélési esélyeit, ezért ezeknél a populációknál az r értéke az evolúció során csökken.
11.4.2. A szimulációs program működése
A program egy Excel táblázat, amely a dN/dt = rN(K-N/K) = rN(1+N/K) képlettel számol. A táblázatban mi adhatjuk meg a kiindulási egyedszám (N1), a szaporodási ráta (r) és a környezet eltartóképességének (K) értékét. Miután ezeket az értékeket megadtuk két egymás alatt elhelyezkedő táblázatban megjelennek az első húsz generáció egyedszámai. Ezeket az adatokat a program a táblázatok alatt szereplő két grafikonon ábrázolja.
Különböző N1 (kezdeti egyedszám), K (környezet eltartóképessége) és r (szaporodási ráta) értékek megadása mellett teszteld a populáció növekedését.
Elsőként az r érték hatását vizsgáljuk meg a grafikon megjelenésében.
Kiindulási adatok:
|
N1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
K |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
|
r |
1,5 |
2 |
2,5 |
3 |
3,5 |
4 |
4,5 |
Elemezd a megjelent grafikonokat!
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
1,5 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
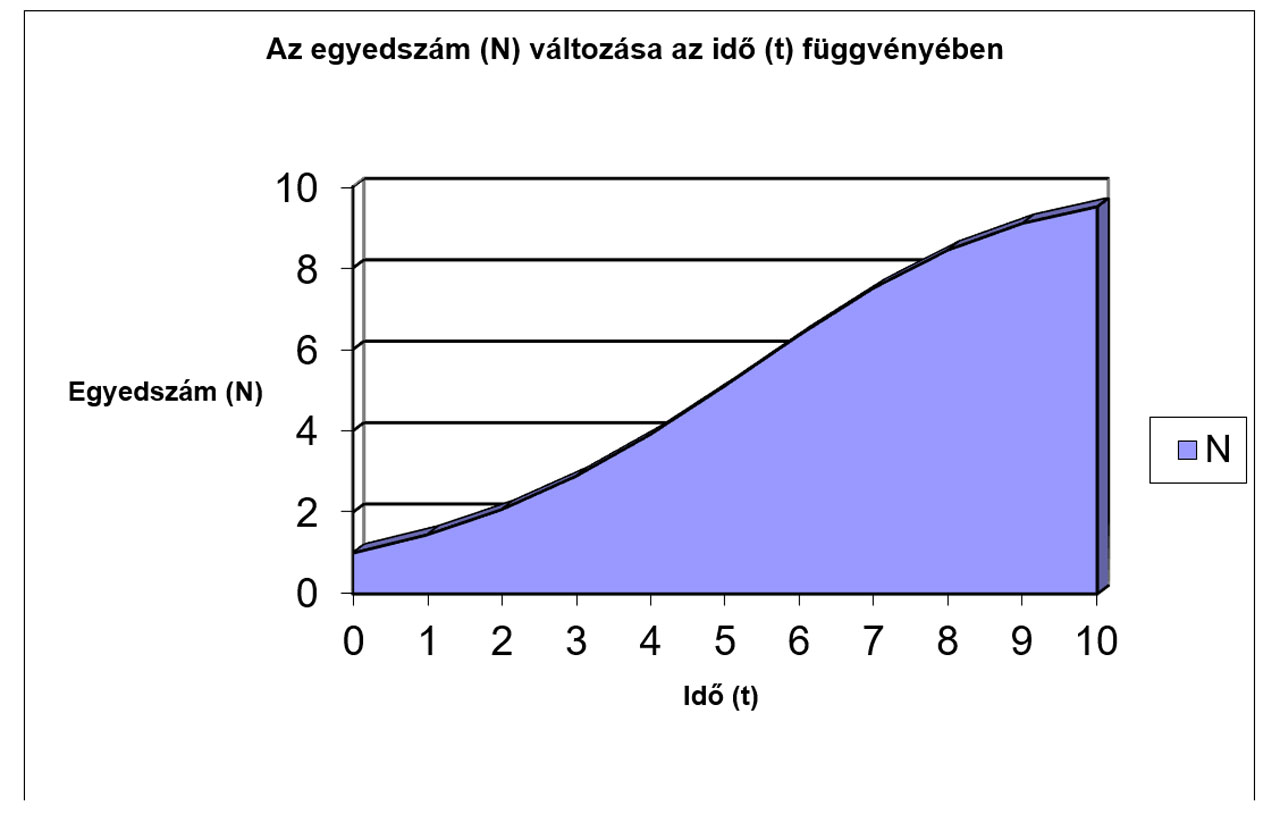
N |
1 |
1,45 |
2,0699 |
2,8906 |
3,9181 |
5,1096 |
6,359 |
7,5166 |
8,45 |
9,1049 |
9,5124 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
N |
9,7443 |
9,8689 |
9,9336 |
9,9666 |
9,9832 |
9,9916 |
9,9958 |
9,9979 |
9,9989 |
9,9995 |
9,9997 |
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
2 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
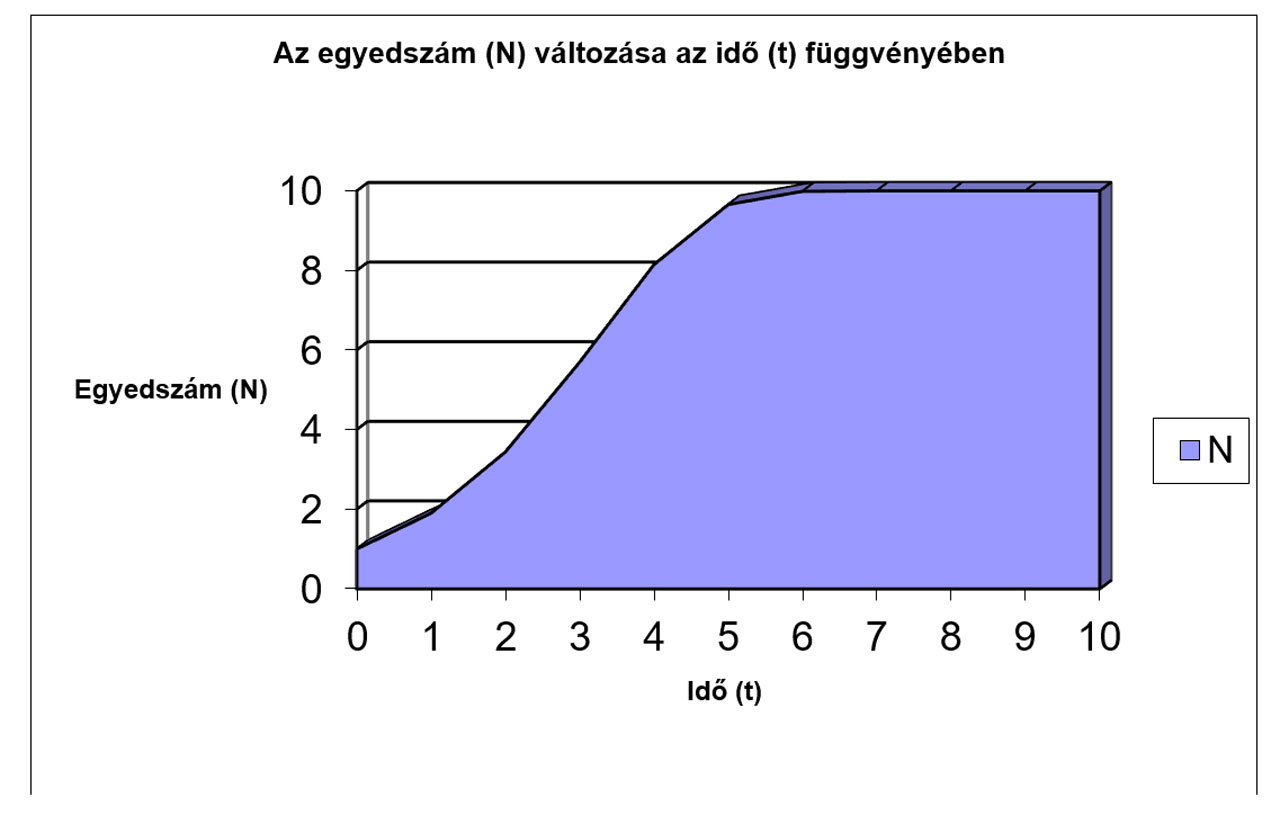
N |
1 |
1,9 |
3,439 |
5,6953 |
8,147 |
9,6566 |
9,9882 |
10 |
10 |
10 |
10 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
N |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
10 |
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
2,5 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
N |
1 |
2,35 |
5,0466 |
8,7963 |
10,385 |
9,7856 |
10,1 |
9,9483 |
10,025 |
9,9872 |
10,006 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
N |
9,9968 |
10,002 |
9,9992 |
10 |
9,9998 |
10 |
10 |
10 |
10 |
10 |
10 |
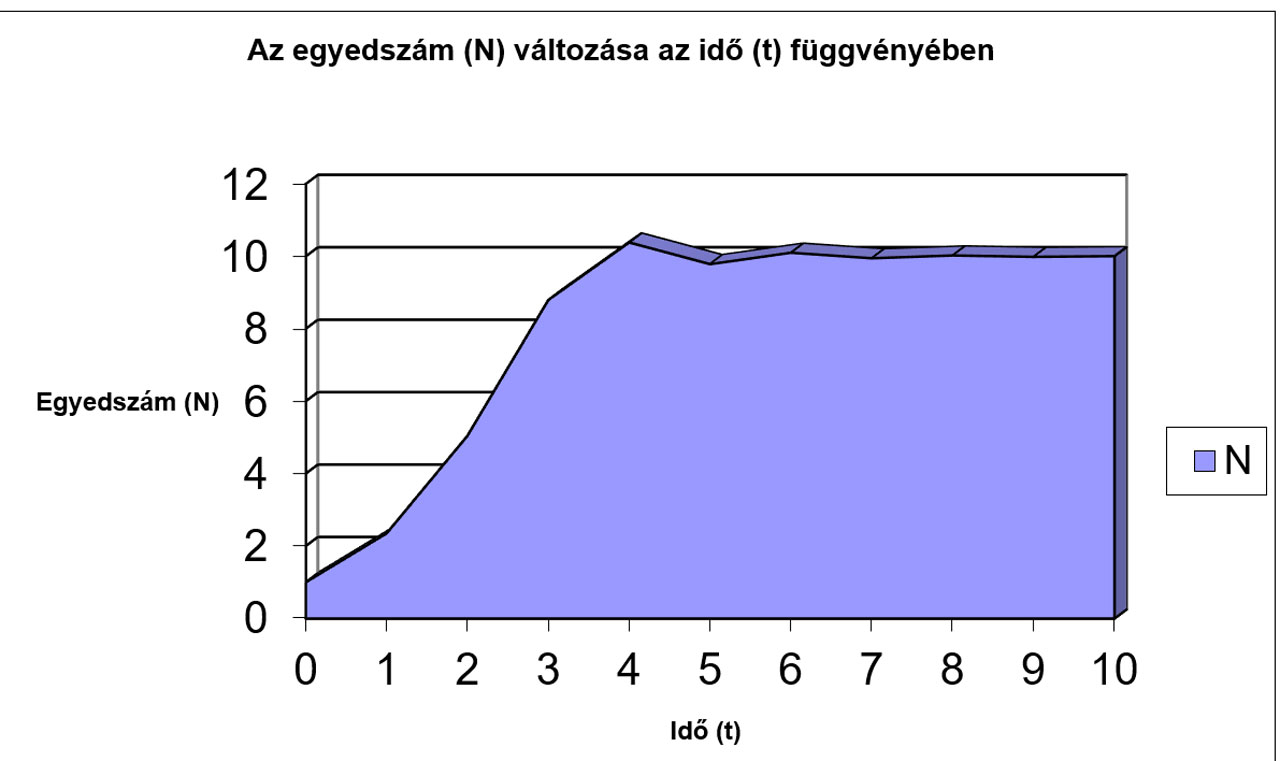
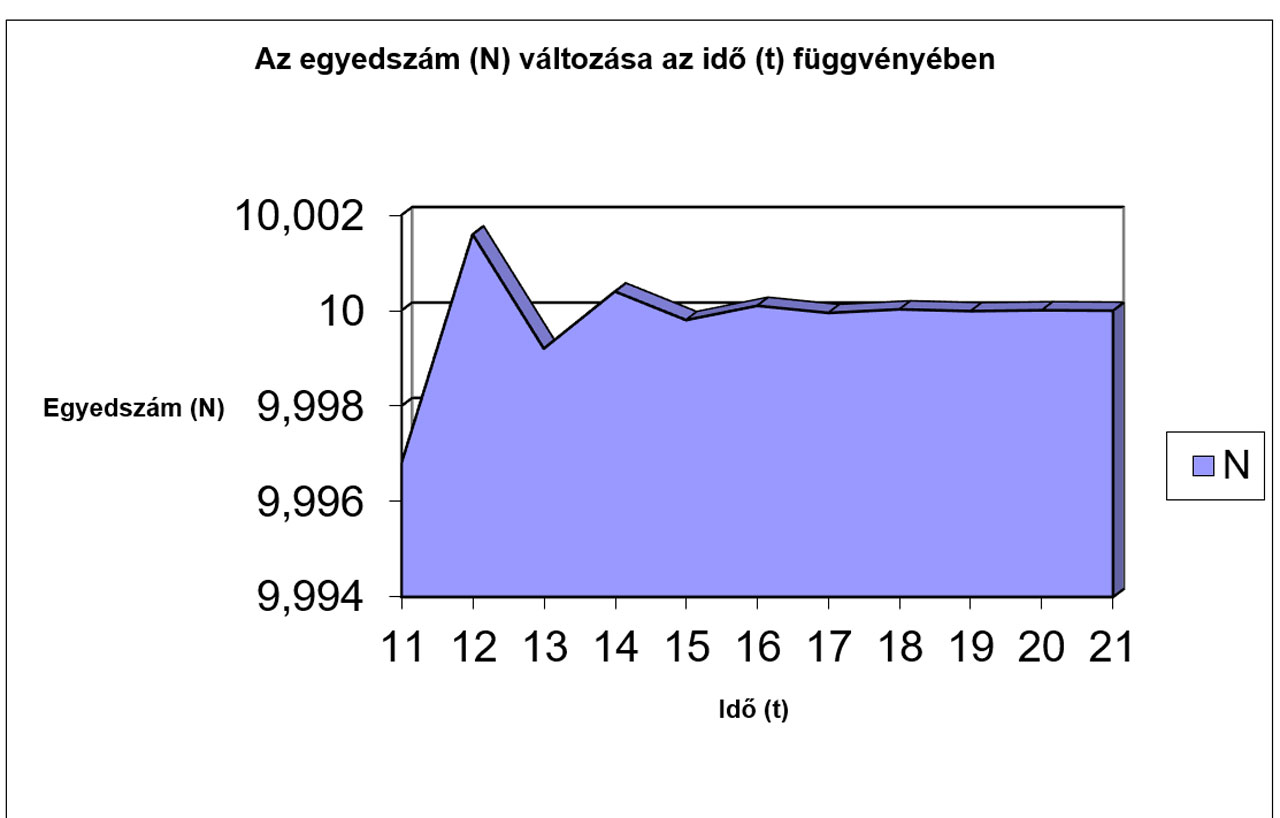
A fenti esetben két grafikonon ábrázoltuk 1-től 21-ig az egyes generációk egyedszámát.
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
3 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
N |
1 |
2,8 |
6,832 |
11,161 |
8,5698 |
11,021 |
8,7703 |
10,927 |
8,9008 |
10,858 |
8,9954 |
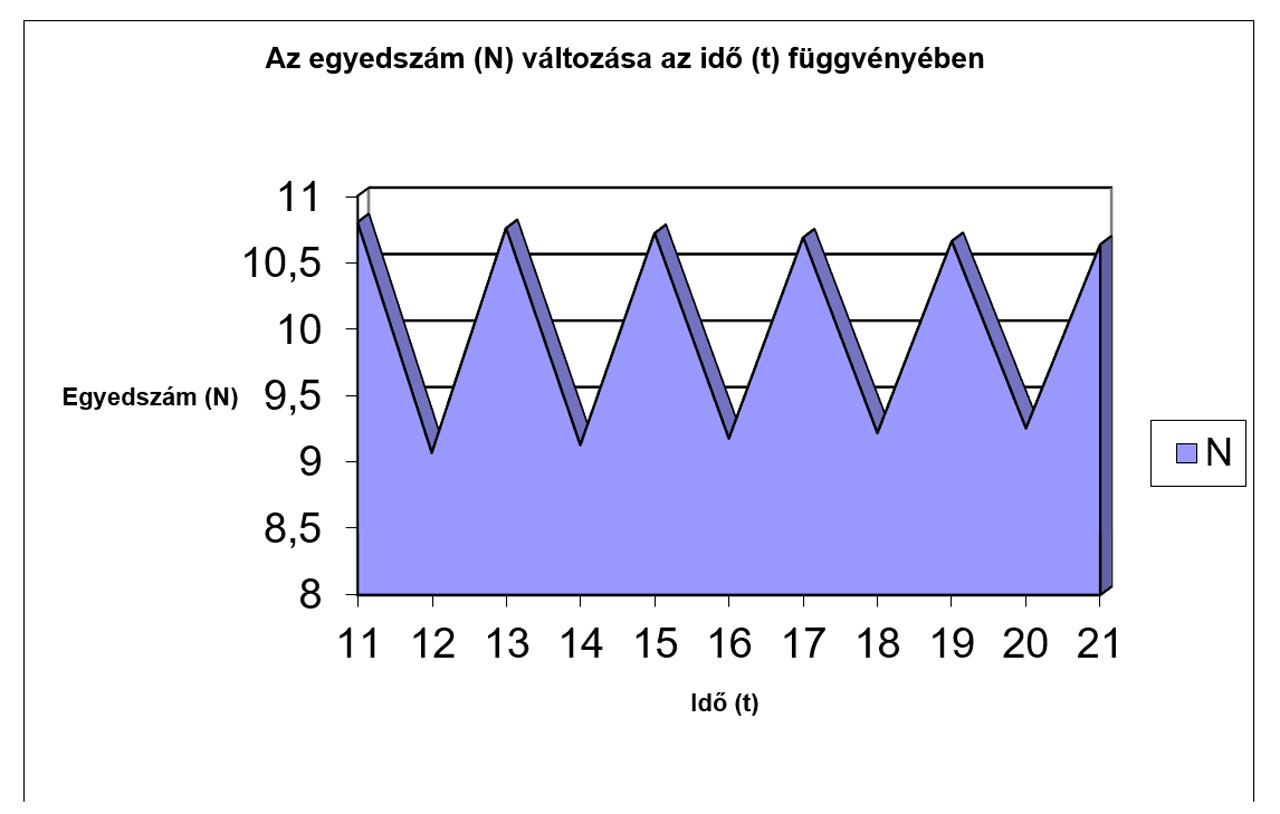
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
N |
10,803 |
9,0683 |
10,758 |
9,127 |
10,721 |
9,1756 |
10,688 |
9,2167 |
10,661 |
9,2521 |
10,636 |
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
3,5 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
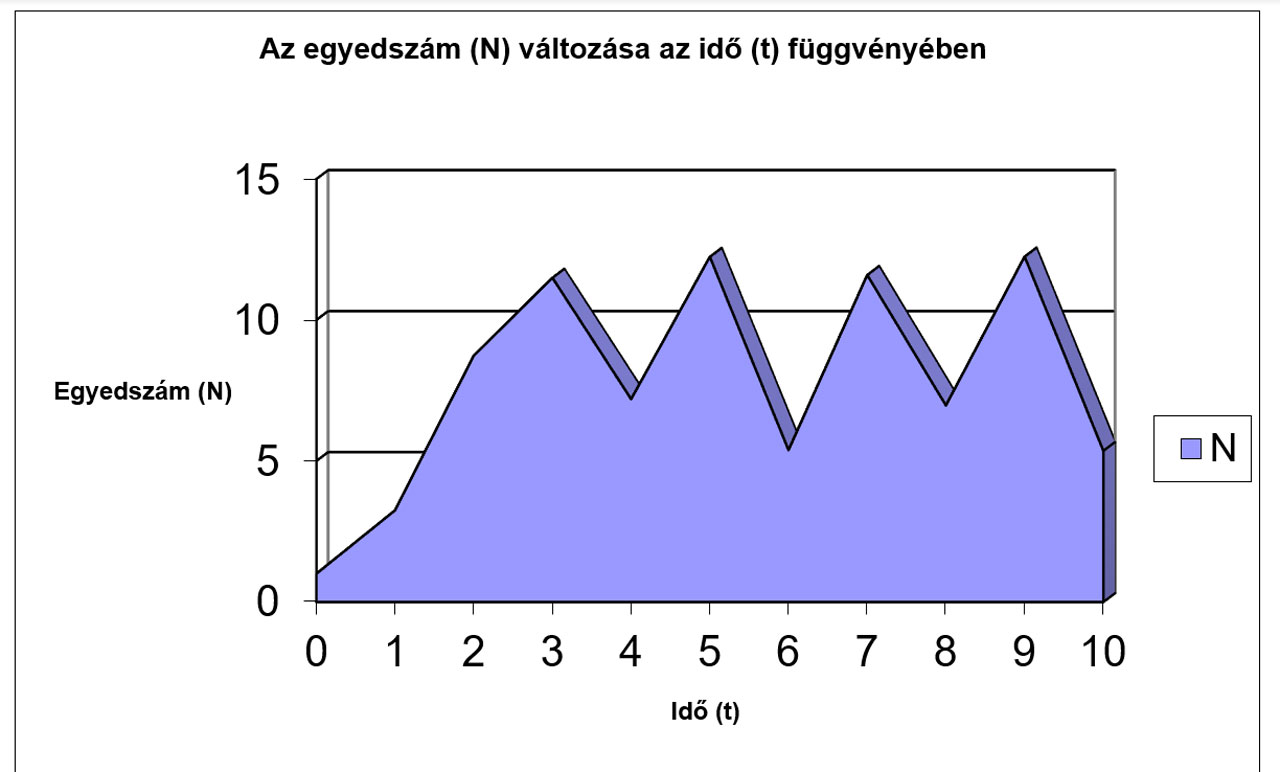
N |
1 |
3,25 |
8,7344 |
11,498 |
7,192 |
12,241 |
5,3836 |
11,597 |
6,9674 |
12,25 |
5,3601 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
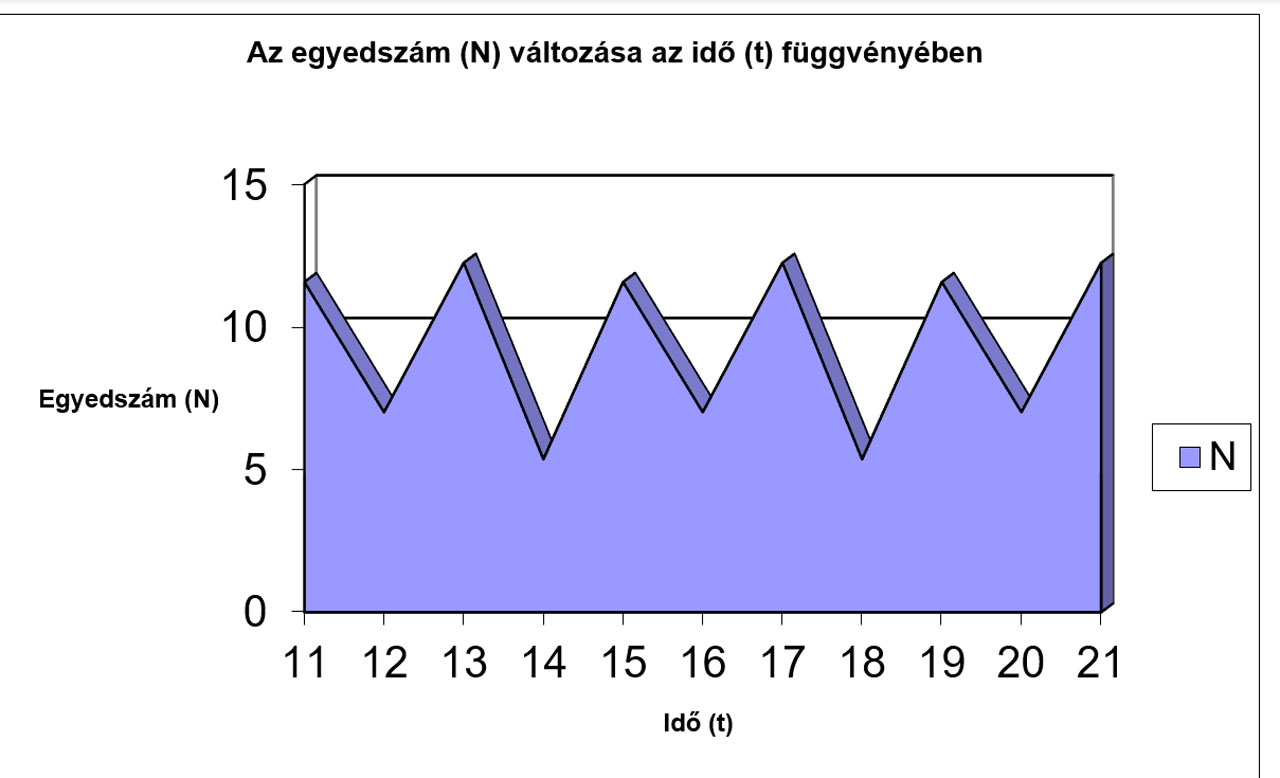
N |
11,578 |
7,0113 |
12,25 |
5,3595 |
11,577 |
7,0124 |
12,25 |
5,3595 |
11,577 |
7,0124 |
12,25 |
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
4 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
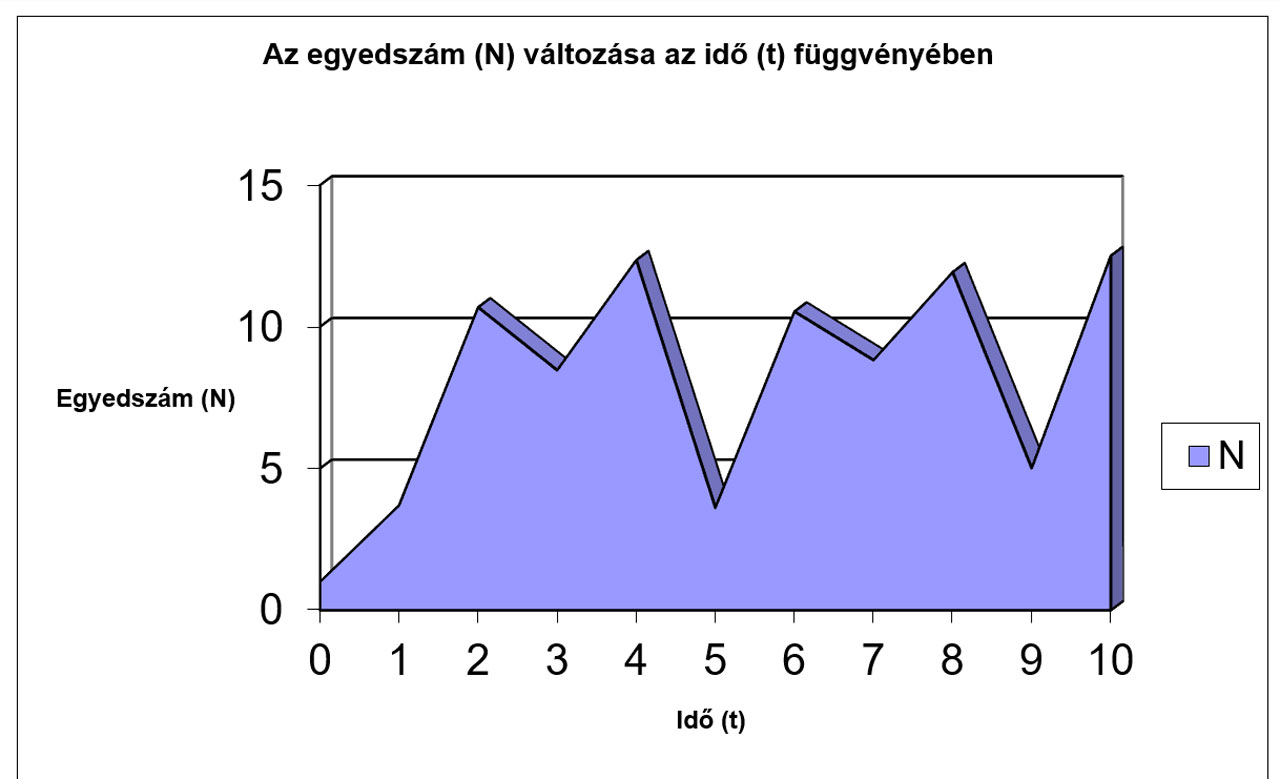
N |
1 |
3,7 |
10,693 |
8,4699 |
12,358 |
3,6166 |
10,542 |
8,8268 |
11,933 |
5,0116 |
12,512 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
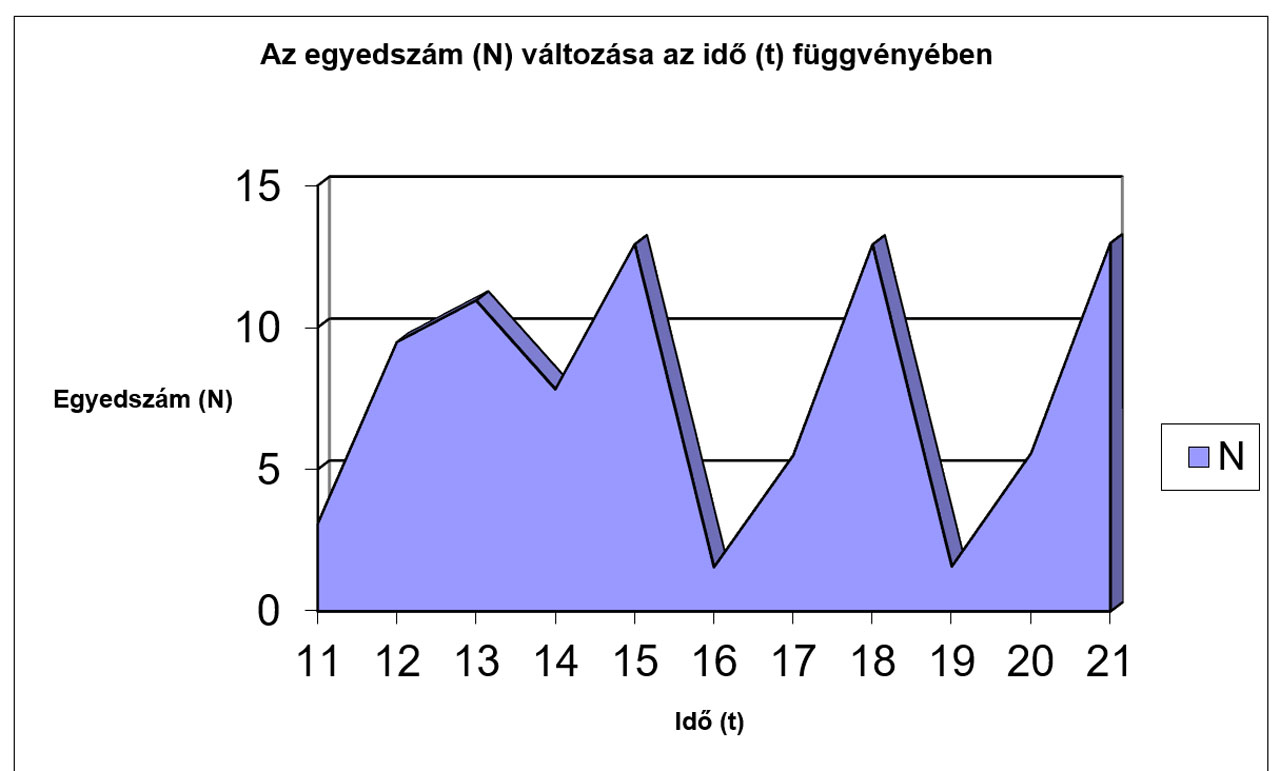
N |
3,0846 |
9,4839 |
10,952 |
7,8233 |
12,932 |
1,5571 |
5,5009 |
12,926 |
1,5809 |
5,5738 |
12,975 |
|
Kiindulási egyedszám, N1: |
1 |
|
|
|
|
|
|
|
|||
|
Környezet eltartóképessége, K: |
10 |
|
|
|
|
|
|
|
|||
|
Szaporodási ráta, r: |
|
4,5 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
N |
1 |
4,15 |
12,647 |
0,9296 |
3,8809 |
12,193 |
2,8359 |
9,9469 |
10,132 |
9,6642 |
10,8 |
|
|
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
|
N |
7,7759 |
13,829 |
-4,704 |
-28,91 |
-422,6 |
-64410 |
-1E+09 |
-7E+17 |
-2E+35 |
-1E+70 |
###### |
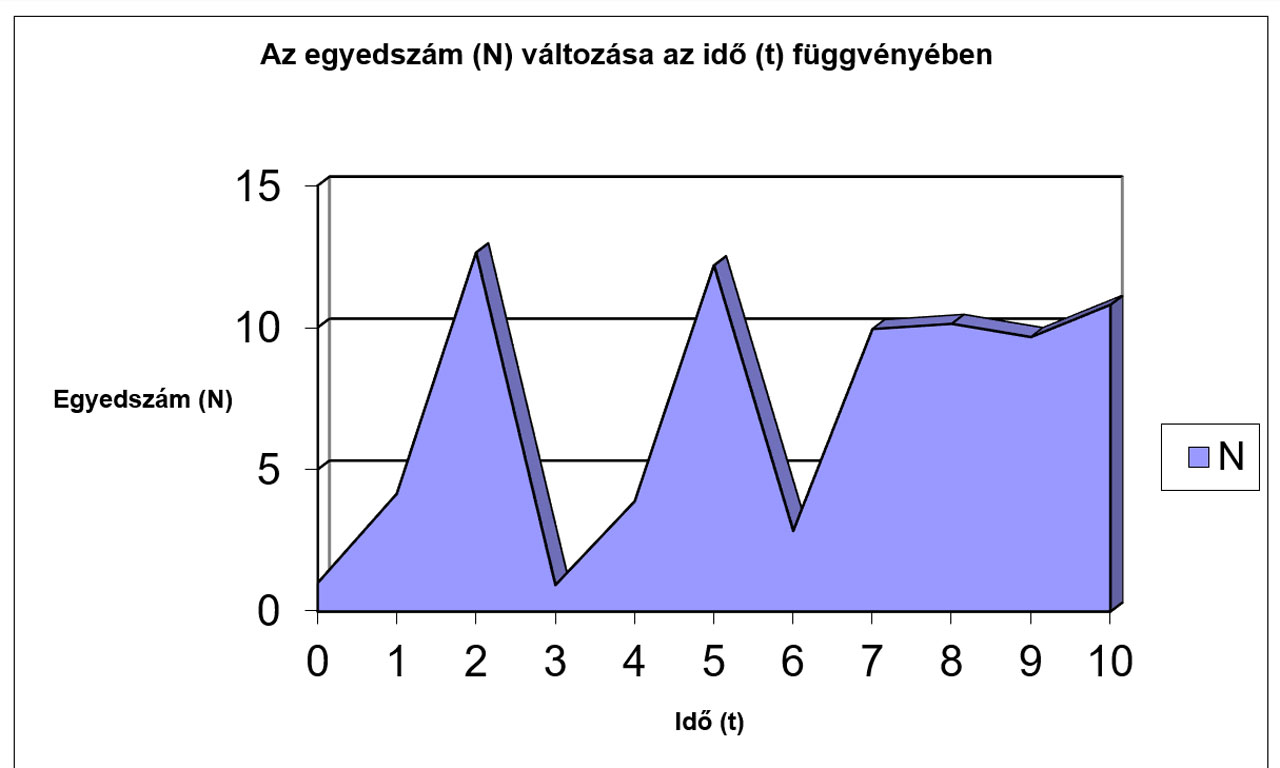
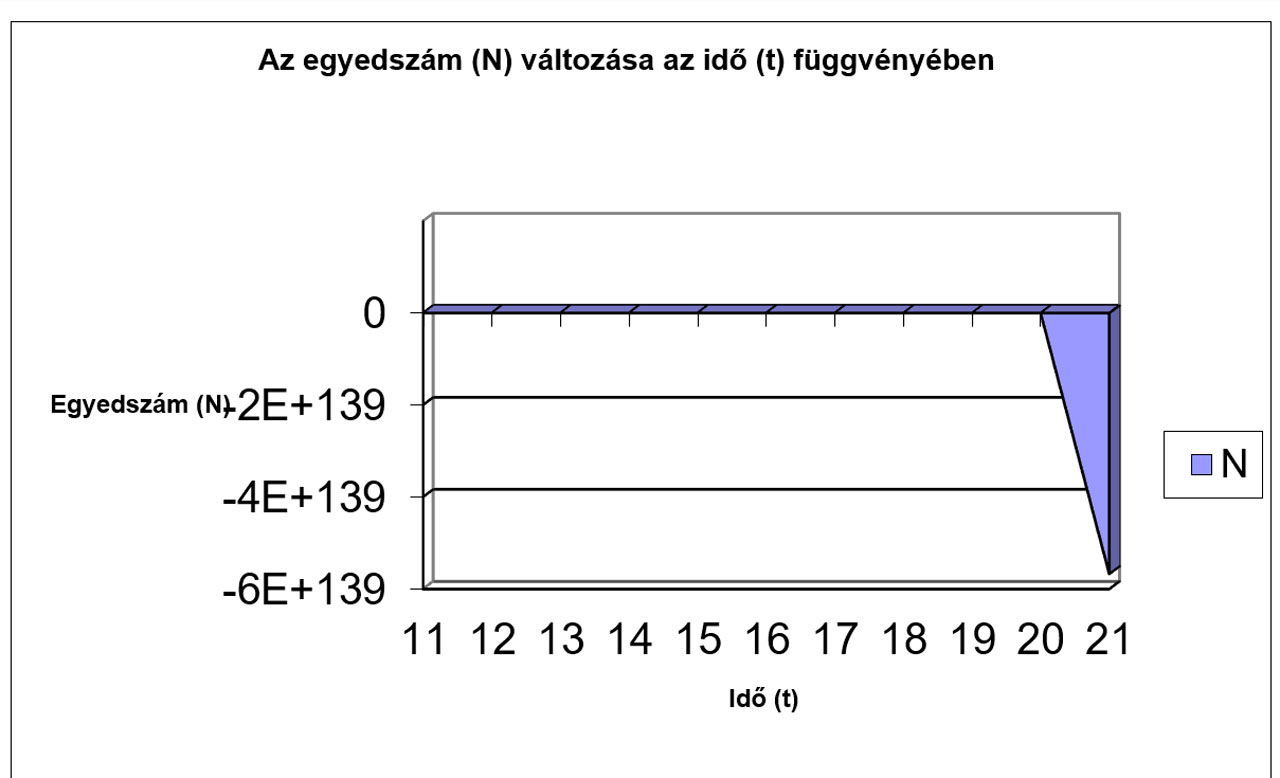
Jól megfigyelhető, hogy az r értékének növekedésével először csak a logisztikus görbe meredeksége változik (a populáció egyedszáma hamarabb éri el a környezet által eltartható értéket (K)), míg később egyre jelentősebbé válnak a túlszaporodásokat követő pusztulások, amik végül oda vezetnek, hogy az egyedszám 0-ra, vagy negatív értékre csökken, ami a populáció kipusztulását jelenti. Az előbbiekhez hasonló módon a K értékének változtatásával szintén tanulságos, sok esetben biológiailag is értékelhető eredményeket kapunk.